
性行为的生理基础
|
[拼音]:xingxingwei de shengli jichu [外文]:physiological basis of sexual behavior 性行为是指动物种内的异性个体之间互相展示的吸引、抚爱和终至于交配的行为。广义的性行为还包括同性个体之间的模拟交配和摆弄自身的生殖器官以满足性欲的行为。 性行为的最终环节是交配。交配行为导致雄性的精子和雌性的卵子相遇。有些动物(如鱼类和两栖类)的两性把精和卵排在体外,使精和卵在水中相遇;而大多数动物则是雄性把精子排入雌性体内,使精卵在雌体内相遇。一个精子钻入一个卵子内,二者的细胞核融合成为受精卵,此后即两个个体的遗传信息共同决定这个新个体的发育。生殖需要两个个体的染色体的新的配合,这一现象在用自身分裂的方式来繁殖的单细胞生物中,如在草履虫中,已经见到。草履虫可以不交配而分裂,在多细胞动物中,也有不经交配而生殖的,称为单性生殖。经雌雄交配生殖的则称为有性生殖。从自然选择的观点来看,有性生殖的优越性在于能增加后代的变异,从而也增加后代的适应性。因此,人们认为有性生殖是自然选择的产物。 在动物界中,性行为的模式是多种多样的。但就雌雄两性的表现形式而言,在各种动物中(除雌雄同体者外)都有明显区别。一般雄性表现主动,雌性表现被动。例如,在鸟类中雄鸟常以各种舞蹈和亮相姿态吸引雌鸟,并以近似狂暴的动作蹬踩雌鸟的背,而雌鸟仅仅以俯伏和撅尾的姿势接受雄性的交配;在哺乳动物中,雄性主动地爬背,雌性以曲背、撅臀和扭尾的姿势接受雄性的交配,这称为接受行为。在高等哺乳动物中,雌性有时也有某种积极求爱的表现,如雌狗在发情时常常爬雄狗的背和嗅雄狗的生殖器,以及在雄狗面前跑来跑去,挑逗雄狗去追逐,这称为前接受行为。人类以下的哺乳动物,交配的姿势大都是雄性在雌性的背方,大猩猩偶尔采取面对面的姿势,人类则以面对面的姿势为主。 两性差异是由个体所具有的染色体决定的。正常人有23对染色体,其中第23对称为性染色体。女性的这对染色体是对称的,通常以XX表示;男性的这对染色体中有一个比较短,故以XY表示。XX或XY染色体只决定最初的性腺的分化。在胚胎的早期(例如人在怀孕第7周之前)性腺是未分化的,这时雌雄两性的表型一样。此后,XX或XY染色体上携带的性别基因开始决定性腺的分化。如果一个个体带有XX染色体,其未分化的性腺的皮质部分开始发育成为卵巢;如果个体的性别染色体是XY,则性腺的髓质部分开始发育成为睾丸。最初的卵巢不分泌雌性激素。而睾丸发育时则分泌雄性激素。 在这以后的发育阶段,有许多实验的事实证明,控制性别分化的主要因素是雄性激素的存在。如果有足够的雄性激素(除睾丸外,肾上腺皮质也分泌性激素)存在,它将促进输精管、贮精囊、阴茎和睾丸的发育。如果不存在雄性激素,或因某种原因使机体对雄性激素不敏感,即使其性染色体是XY型,它的生殖系统也不能发育成正常雄性的样子。带有XX染色体的雌性,在发育中没有雄性激素的影响通常都能发育成为正常的雌性,有发育的卵巢、子宫和阴道。但是,如果在发育过程中由于某种原因(例如母亲怀孕或哺乳时期服用大量含雄性激素的药物)受到了雄性激素的影响,其雌性生殖系统的发育就会受到抑制,甚至于外生殖器长成雄性的样子。 性激素不仅在个体发育的早期决定两性生殖系统的分化,而且也决定成熟个体的性行为的性别差异。给一个刚出生的雌仓鼠注射大量(100微克)睾丸酮,它在成熟之后会表现对雌仓鼠爬背的雄性行为。再给它注射雌激素和黄体酮,它也不会再表现出雌性的性接受行为。通常在实验室中都可以观察到,阉割了的雄性大鼠和家兔对发情的雌性都不表现雄性的爬背行为,但给它们注射睾丸酮后,可以使它们正常的雄性性行为得到恢复。此外,在人类以下的哺乳动物中,雌性有受体内雌性激素水平控制的发情周期,雄性则因雄性激素对下丘脑的作用,体内既无性激素水平的变化,行为上也不表现周期性的发情。 性激素对成年个体性行为的影响,在不同种类的动物中程度是不一样的,雄性大鼠和家兔阉割之后性行为即消失,雄狗阉割后需要几个月性行为才逐渐衰退,恒河猴的这一过程要一年多。在成人,如果由于某种原因失去了睾丸,有的要经过几年才丧失性欲和性兴奋。有过性经验的成年妇女在作过卵巢切除手术之后,多数都不影响性生活。这是因为控制性兴奋和性行为的神经机制是多层次的。应该说,随着大脑皮层的发育,控制性兴奋的因素越来越复杂化(见图)。 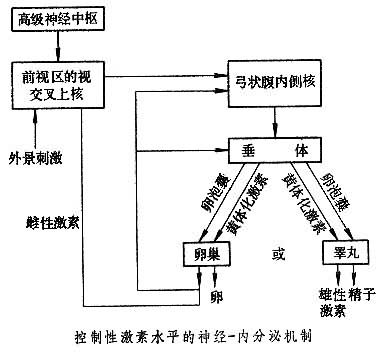
控制反射性的性兴奋(例如用手按摩阴茎而引起的阴茎勃起和射精)的中枢在脊髓的腰腱部分。这一低级中枢的兴奋,可以由高级中枢的活动促进或抑制。但在脊髓低部位切断之后,阴茎局部的适当的触觉刺激也可以引起阴茎勃起和射精。 受激素的影响而控制性行为的中枢主要部分在下丘脑。在雄性中,下丘脑的内侧前视区和与其连接着的内侧前脑束是控制雄性性行为的重要结构。电刺激雄大鼠的这部分,可促进其交配活动。将微量睾丸酮置于阉割的雄大鼠的这一区域可使它更多地从事性行为。切断这一区域和内侧前脑束的连接则使雄性大鼠的交配和射精发生困难。主要原因可能是对引起性兴奋的某些刺激的反应力降低。内侧前视区对于雌性表现雄性的爬背行为是重要的。但对于雌性表现性接受行为是不重要的。对于雌性表现性接受行为的重要区域是前下丘脑区。损伤此区之后,雌大鼠即不再表现脊柱前曲性接受行为。将雌性激素置于此处可使雌大鼠出现性接受行为。此外,中脑网状结构似亦参与雌大鼠的性行为,因此如将黄体酮置于一个预先用过雌激素的雄大鼠中可以促进它的性接受行为。 在人类,有事例证明,下丘脑的腹内侧核经手术破坏后可使男性的性行为消失。 在这些中枢之上,还有更高级的控制性行为的中枢,包括边缘系统中的杏仁核、海马、颞叶和额叶腹内侧的部分皮层。这些中枢中介来自外部世界的刺激(如周围环境的条件、异性对象的某些躯体的和行为的特点以及与习得的认识和情绪有联系的某些特殊刺激)对性行为的影响。事实上,越是高等的动物,其性行为越是受高级中枢的控制。例如,性激素在控制鼠和兔的性行为中作用十分明显,但在有过性经验的雄狗和雄猴中,阉割后(取消雄激素)性兴奋的能力还保持相当长的时间。在有过性经验的男人和女人中,性激素的水平对性行为的影响更不明显。 性行为在维持种族的生存和繁衍中虽然和饮食行为同等重要,但也有其不同之处,最重要的不同是不饮不食个体是一定要死亡的,没有性的满足个体仍能生活。此外有效的性行为必然要有异性个体的配合才能完成。由于它涉及到两个个体的关系,在文明的人类社会中,这种关系又牵扯到许多复杂的社会问题,所以它受许许多多社会条件的制约。诸如风俗习惯、道德观念以及个人在生活中习得的一些特殊的好恶和对性的态度等都影响性的欲求和性行为的表现。可以认为,这些复杂的关系,通过人的大脑皮质的活动,在调整人的性行为中起着独特的作用,而性激素的影响则退居相当次要的地位,而且这种影响也受高级中枢活动的控制。
|
















