
染色体
|
[拼音]:ranseti [外文]:chromosome 染色体词条主要包括发展简史、数目和大小、数目、大小、化学组成、DNA、组蛋白、非组蛋白、中期染色体的形态、着丝粒、核仁组织区(NOR)、端粒、随体、染色体的结构、核小体、染色体的高层次结构、常染色质与异染色质、结构异染色质、兼性异染色质、性染色体与性别、B染色体、染色体的复制、理论和实践意义、染色体的遗传学说、临床医学、环境科学、农业生产等方面的知识。 真核细胞有丝分裂和减数分裂时由染色质聚缩而成的结构,一般呈棒状,因易被碱性染料染色故称染色体。染色质是由核内的脱氧核糖核酸(DNA)与组蛋白、 非组蛋白等结合形成的线状结构。染色质在有丝分裂之前即开始聚缩,至分裂中期呈现为具明显种属特征的染色体。染色质和染色体是在细胞周期不同阶段可以互相转变的形态结构。 除去一般形态的染色体外,在一些生物的卵母细胞等细胞中还出现灯刷染色体,在双翅目昆虫的某些细胞中出现多线染色体。 真核细胞的绝大部分DNA集中在染色体中,而DNA记载着遗传密码,所以染色体是细胞中主宰遗传的结构。 原核细胞的DNA也常被称为染色体。但它的DNA并不象真核细胞的染色质那样与组蛋白等形成复合体,在细胞分裂时也不聚缩。 研究简史1848年德国植物学家W.霍夫迈斯特在紫露草属小孢子母细胞中最早看到相当于染色体的物体。1875年德国动物学家O.赫特维希看到受精时两个配子融合后着色物体数目加倍,这一事实不久得到E.van贝内登等人的证实。W.弗勒明1880年发现有丝分裂时每条着色物体都纵裂为二,并分别进入两个子细胞。1882年他提出在同一物种的细胞中着色物体数目是恒定的。1888年W.瓦尔代尔正式提出染色体的命名。 1900年重新发现G.J.孟德尔的遗传法则后不久,W.S.萨顿和T.H.博韦里分别提出在减数分裂过程中染色体的行为与G.J.孟德尔设想的遗传因子的行为之间有平行关系,因此认为遗传因子存在于染色体上。这个假设1910年以后被T.H.摩尔根等证实并发展。摩尔根等证明:遗传因子(基因)在染色体上呈线形排列。 1924年R.孚尔根等制定DNA的组织化学反应法,后来T.O.卡斯珀松又建立测量 DNA的紫外线显微分光光度法。用这些方法都发现 DNA存在于间期核和染色体中。O.T.埃弗里1944年发现细菌转化因子是DNA,首次证实DNA是遗传物质,从而找到了染色体做为遗传结构的分子基础。 1953年J.D.沃森和F.H.C.克里克提出了 DNA双螺旋分子结构模型,对 DNA的认识取得突破性进展。1957年J.H.泰勒等,1958年M.S.梅塞尔森等先后证明染色体DNA的复制是半保留性的。这就说明染色体和脱氧核糖核酸确有作为遗传结构应该具备的自我复制功能。 从50年代末到60年代对染色质和染色体的超微结构提出过一些设想的模型,但都因缺乏根据而未能得到肯定。1973年A.L.奥林斯等在间期核中发现核小体,并通过许多学者的研究对核小体分子水平的结构确立了大体相似的模型。现已公认核小体是染色质的基本单位(亚单位)。至于由核小体到染色体的高层次结构还正在探索。 数目和大小数目各种生物的染色体数目不同。大多数生物的体细胞含有12~50个染色体(2n)。少的如植物的纤细单冠菊和动物的壳虫都只有4条染色体;最多的是一种瓶尔小草(Ophioglossum reticulatum)2n=1260和一种蝴蝶(Lymandra atlantica)2n=446。人的染色体数为46,牛38,猪40,狗78;普通小麦为42,水稻24,玉米20,葱16,百合24。 植物中染色体数目常呈倍性现象。例如小麦属的一粒小麦染色体数为14,二粒小麦为28,普通小麦为42。就所含基本染色体组而言它们分别为二倍体、四倍体和六倍体植物(见染色体倍性)。 大小各种生物的染色体大小不一,长度以1~10微米最为常见,直径多为1~2微米。通常植物的比动物大。植物中染色体较大的如直立延龄草;较小的如加州藨草约有100多个微小染色体;动物中以蝗虫、螳螂和蝾螈的染色体较大,鸟类和爬行类除有较大染色体外常含有许多微小染色体。 化学组成中期染色体与间期染色质的化学组成相似,都含有DNA、组蛋白、非组蛋白和核糖核酸(RNA)以及少量脂类、钙和镁(见表)。 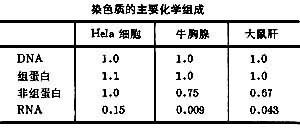
DNA不同生物的单倍体细胞中DNA含量不同,在同一种生物的单倍体细胞中DNA含量一般是恒定的;高等生物细胞核内 DNA含量比低等生物的高,但也有相反现象。在亲缘关系近的种属间,细胞核内DNA含量也可相差很大。 组蛋白属碱性蛋白,主要有5种(H1、H2A、H2B、H3和H4)。组蛋白缺乏组织专一性,种属之间差异少。 非组蛋白至少包括50~100种不同的蛋白质,其中80%为酸性蛋白,具有种属专一性。有人认为非组蛋白能专一地激活某些基因。 中期染色体的形态中期染色体具有比较稳定的形态。它由两条相同的姐妹染色单体构成,并在纵轴方向呈现一些变细的部位(图1);分别称为主缢痕和次缢痕。前者是着丝粒的所在地,后者为核仁组织区的部位。 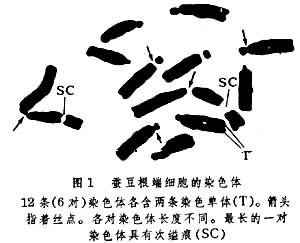
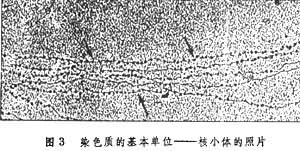
着丝粒在主缢痕区存在着丝粒,由此把染色体分成二臂。着丝粒的两侧各有一由蛋白质构成的三层的盘状或球状结构,称着丝点,着丝点与纺锤体的纺锤丝连接,与染色体的移动有关。在分裂的前期和中期,着丝粒把两个姐妹染色体连结在一起,到后期,两个单体的着丝粒分开,纺锤丝把两条染色单体拉向两极。 根据着丝粒在染色体上的不同位置,可以分为中间着丝粒染色体、近中着丝粒染色体、具近端着丝粒染色体和端着丝粒染色体4种。 核仁组织区(NOR)存在于某一染色体的次缢痕区。在间期核中由染色体的这一区域形成核仁,故名。核仁组织区是由核糖体 RNA基因构成的。这种基因转录形成rRNA,而rRNA的积累和发育,使核仁增大。通常每组染色体中至少有一个NOR。 端粒指染色体末端部分的特殊构造。它不呈现特殊的形状,但可起防止染色体末端彼此粘连的作用。染色体在丢失端粒后易与其他染色体末端互相粘连,或与同一染色体的另一端连接而成环形。 随体染色体末端部分的粒状结构,通过次缢痕区与染色体的主体部分相联。它是识别染色体的重要特征之一。 染色体的结构核小体构成染色体(染色质)的基本单位。它的核心是由4种组蛋白(H2A、H2B、H3和H4)各2分子组成的八聚体,外绕DNA分子(大约146碱基对,围绕1.75周),称为核小体核心颗粒(图2)。完整的核小体为 110×110×57埃的扁圆柱状小体,DNA在组蛋白八聚体外缠绕2周(约166碱基对)。核小体靠DNA互相连结成串珠状结构(图3)。连结两个相邻核小体的 DNA长度随生理状态的不同而异,一般为60碱基对。靠近每个核小体结合着一分子的组蛋白H1。按R.D.昆博格和A.克鲁格的模型,H1位于DNA超螺旋进出核小体区的一侧。 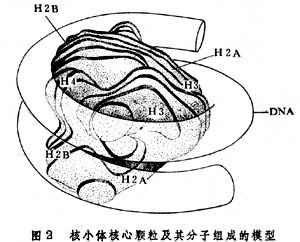
染色体的高层次结构每一染色单体由一条 DNA蛋白纤维盘绕而成。因为从酵母或黑腹果蝇细胞核内抽提的DNA分子大小与一条染色体的DNA平均含量相近。由核小体串连成的细线称核丝,直径约100埃。由核丝如何聚缩成染色体有各种假说,尚无定论。按多级螺旋假说;核丝螺旋成直径约 300埃的染色纤维,称螺线管,后者再螺旋成直径为2000~4000埃的染色线,称超螺线管。这种染色线再螺旋乃成直径约 1~2微米的染色体(图4)。 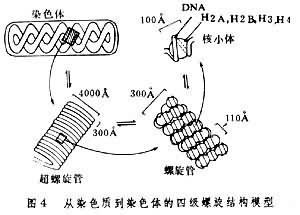
按有的模型,由300埃的染色质纤维形成染色体的过程不是靠螺旋,而是靠随机折叠或形成侧环。不同的模型都有或多或少的事实根据。 常染色质与异染色质在细胞周期中显示正常周期变化的染色质称为常染色质;同一染色体的各段落或片段,其凝缩-松散的交替变化不一致,在间期和前期,染色体过度凝缩和超螺旋化,因而染色很深,或者在中期染色体凝缩不足和局部解旋,因而染色很浅,这些变化不正常的染色体片段,统称异染色质。在DNA合成期异染色质区比常染色质区晚复制。异染色质分结构异染色质和兼性异染色质两大类。 结构异染色质间期细胞核内经常呈现浓集的染色质团块,通常多位于核膜下,其中含有高度重复顺序的DNA;常分布于染色体的着丝粒区、 端粒和次缢痕附近。核内异染色质团块有时称为前染色体或染色中心。每一团块代表一个染色体的着丝粒区异染色质。此外,在间期细胞核内包围核仁的凝缩团块特称与核仁结合的异染色质。 兼性异染色质某些生物在一定发育时期的间期核内,原来的常染色质凝缩,并丧失基因活性,变为异染色质,称之为兼性异染色质。例如雄性哺乳动物的单个X染色体呈常染色质状态,而雌性哺乳动物体细胞核内,两个X染色体之一异染色质化。在上皮细胞核内,这个异固缩的X染色体称性染色质或巴氏小体。在多型核白细胞核内,此X染色体形成特殊的“鼓槌”结构。雄性动物无此特征。因此检查羊水中胚胎细胞的巴氏体可以预测胎儿的性别。 性染色体与性别在雌雄异株植物和雌雄异体动物中,决定性别或影响性器官分化的染色体称性染色体,其余为常染色体。动物的性染色体有三种类型: (1)XX(雌),XY(雄)型,例如,哺乳动物雄性个体有两个不同的性染色体(X、Y),故能产生两种不同的配子。一种带X染色体,一种带Y染色体。但雌性的两个性染色体是同源的(X、X),只能产生相同的配子。受精时带X的精子与卵子结合产生雌性个体,而带Y的精子与卵子结合产生雄性个体。 (2)ZW(或XY)(雌)、ZZ(或XX)(雄)型,例如鸟类和爬行类等,雌性产生含Z或W两种配子,雄性产生含Z配子。 (3)XX(雌),XO(雄)型,例如某些昆虫,雌性产生含X的一种配子,雄性产生含X或不含性染色体的配子。 在苔藓植物有些雌雄异株的单倍体配子体中,有X染色体的为雌性,有Y染色体的为雄性,含有任何比例的X和Y为雌雄同株,而且不育,但双倍体孢子体(XY)无性别区分。许多雌雄异株的被子植物也有性染色体。例如杨树和柳树的性别都是由性染色体决定的。它们的染色体也是XX(雌)和XY(雄)类型。 B染色体细胞内有一定数目、形态和结构的染色体称 A染色体,包括常染色体和性染色体在内。它们在遗传上是重要的,对个体的正常生活和繁殖是必需的。其数目的增减和结构的变化对机体会造成严重后果。但是,某些生物细胞中可见超数染色体称为B染色体。在演化上B染色体系由A染色体衍生,多为遗传惰性,对个体的表型无多大影响。 B 染色体在植物中较动物更为常见,前者如在玉米、野百合、大花延龄草和黑麦等,后者如在蜗牛、蝗虫、甲虫和介壳虫、以及除鱼和灵长类以外的脊椎动物。 B染色体通常比A染色体小,它们互相不同源,在减数分裂时不相互配对。即使配对也无交叉。B 染色体大部分为异染色质,但也有部分或全部为常染色质的。每个细胞有1~2个B染色体对个体的表型无大影响,在数目较多的情况下则会影响个体生活力和生育力。 染色体的复制复制是由一条变成两条在形态结构、基因排列和分子组成上都完全相同的染色体的过程。染色质复制包括脱氧核糖核酸的复制、染色质蛋白质的合成以及由 DNA和组蛋白形成染色质的基本单位──核小体的过程。 DNA在S期复制(见细胞周期),它的两条多核苷酸链在一些部位逐渐分开,按核苷酸碱基互补配对原则(A-T;G-C),以原来的两条多核苷酸链为模板,复制成两条完全相同的DNA分子。 DNA的复制并不是由一端到另一端连续进行的,而是存在着分成许多小区段的复制单位。这种复制单位亦称复制子。每个复制单位由一点开始向相反的两侧推进,至与相邻的复制单位相遇而结束复制的过程(图5)。 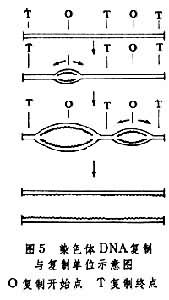
染色体DNA的各复制单位并不都是一同开始复制的,而是按一定顺序有规律地进行复制。通常是常染色质的DNA先复制,而异染色质区的DNA后复制。 组蛋白在细胞质的多核糖体上合成,和细胞核内DNA的复制大体上同时进行。合成的组蛋白迅速输送到核内,所以细胞质中的组蛋白贮存很少。随着 DNA复制的进行,输入的组蛋白很快就和它组装成核小体结构(图6)。 
复制后的染色质经过螺旋化或折叠过程,在有丝分裂时形成具有两条染色单体的染色体。在有丝分裂后期和末期,两条染色单体分到两个子细胞中去。 理论和实践意义染色体的遗传学说遗传定律与染色体行为之间的平行关系证明染色体是遗传基因的载体。如孟德尔的独立分配定律的基础在于减数分裂过程中,配对的同源染色体彼此分离、各趋一极;自由组合定律则在于减数分裂中,同源染色体的自由分配、非同源染色体的随机组合,以及不同雌雄配子在受精时的组合。摩尔根的连锁定律则由于同一染色体内的遗传物质一同传递的结果。因此,基因连锁群数与单倍体细胞的染色体数目相等。互换定律则是基于非姐妹染色单体之间交换染色体片段。从基因交换频数与基因位置之间的距离成正相关,推测基因在染色体上成线性排列。从基因DNA转录出RNA指导专一蛋白质和酶的合成的事实,确认基因可以决定个体的形态结构和功能等表型,控制个体的发育过程。因此,对染色体的研究已成为探索生命本质的重要课题。 临床医学染色体病和肿瘤细胞都伴有染色体畸变。畸变既涉及发病原因又是诊断的依据。婚前检查夫妇,或产前检查胎儿是否存在染色体畸变,是推行优生的重要依据。 环境科学应用检测染色体断裂和姊妹染色单体互换 (SCE)等技术可推断环境中诱变剂和电离辐射的毒性程度,从而采取措施保护人类健康。 农业生产单倍体花粉培养、异源多倍体和染色体工程等技术可用于改良作物品种和培育新种。
|



















